Abstract
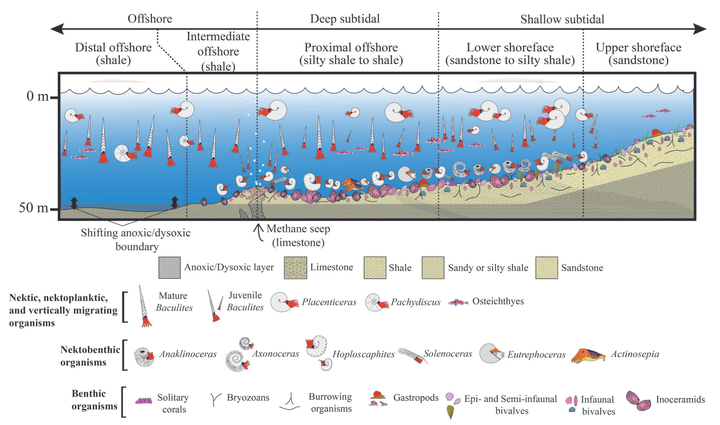
Heteromorphs are ammonoids forming a conch with detached whorls (open coiling) or non-planispiral coiling. Such aberrant forms appeared convergently four times within this extinct group of cephalopods. Since Wiedmann’s seminal paper in this journal, the palaeobiology of heteromorphs has advanced substantially. Combining direct evidence from their fossil record, indirect insights from phylogenetic bracketing, and physical as well as virtual models, we reach an improved understanding of heteromorph ammonoid palaeobiology. Their anatomy, buoyancy, locomotion, predators, diet, palaeoecology, and extinction are discussed. Based on phylogenetic bracketing with nautiloids and coleoids, heteromorphs like other ammonoids had 10 arms, a well-developed brain, lens eyes, a buccal mass with a radula and a smaller upper as well as a larger lower jaw, and ammonia in their soft tissue. Heteromorphs likely lacked arm suckers, hooks, tentacles, a hood, and an ink sac. All Cretaceous heteromorphs share an aptychus-type lower jaw with a lamellar calcitic covering. Differences in radular tooth morphology and size in heteromorphs suggest a microphagous diet. Stomach contents of heteromorphs comprise planktic crustaceans, gastropods, and crinoids, suggesting a zooplanktic diet. Forms with a U-shaped body chamber (ancylocone) are regarded as suspension feeders, whereas orthoconic forms additionally might have consumed benthic prey. Heteromorphs could achieve near-neutral buoyancy regardless of conch shape or ontogeny. Orthoconic heteromorphs likely had a vertical orientation, whereas ancylocone heteromorphs had a near-horizontal aperture pointing upwards. Heteromorphs with a U-shaped body chamber are more stable hydrodynamically than modern Nautilus and were unable substantially to modify their orientation by active locomotion, i.e. they had no or limited access to benthic prey at adulthood. Pathologies reported for heteromorphs were likely inflicted by crustaceans, fish, marine reptiles, and other cephalopods. Pathologies on Ptychoceras corroborates an external shell and rejects the endocochleate hypothesis. Devonian, Triassic, and Jurassic heteromorphs had a preference for deep-subtidal to offshore facies but are rare in shallow-subtidal, slope, and bathyal facies. Early Cretaceous heteromorphs preferred deep-subtidal to bathyal facies. Late Cretaceous heteromorphs are common in shallow-subtidal to offshore facies. Oxygen isotope data suggest rapid growth and a demersal habitat for adult Discoscaphites and Baculites. A benthic embryonic stage, planktic hatchlings, and a habitat change after one whorl is proposed for Hoploscaphites. Carbon isotope data indicate that some Baculites lived throughout their lives at cold seeps. Adaptation to a planktic life habit potentially drove selection towards smaller hatchlings, implying high fecundity and an ecological role of the hatchlings as micro- and mesoplankton. The Chicxulub impact at the Cretaceous/Paleogene (K/Pg) boundary 66 million years ago is the likely trigger for the extinction of ammonoids. Ammonoids likely persisted after this event for 40–500 thousand years and are exclusively represented by heteromorphs. The ammonoid extinction is linked to their small hatchling sizes, planktotrophic diets, and higher metabolic rates than in nautilids, which survived the K/Pg mass extinction event.
Benjamin J. Linzmeier
Assistant Professor of Earth Sciences
My research interests include biomineralization and geochemistry of sedimentary rocks and fossils.